Wide Crosses Among Succulent Plants
Text & Photo: Gordon D. Rowley
Cactus & Co. (Italy) 4(6) 2002

Title page |
Succulents and orchids stand apart from most other flowering plants in the
ease with which wide crosses can be made. A wide cross is a hybrid between
two plants so dissimilar in appearance that one would not expect them to
interbreed. Since classical times it has been recognised that it is rarely
possible to hybridise animals or plants that are not obviously closely
related. A rose will cross with another rose, but not with a peony or a
camellia. But among the succulents we encounter many surprises.
Both succulents and orchids are highly specialised in evolutionary terms,
but it seems that the basic flower structure has diverged less than the
plant body, at least in setting up genetical barriers to outcrossing that
usually accompany (and initiate) speciation. Under favourable conditions in
cultivation, when plants widely separated in the wild are brought together
in one glasshouse, remarkable intermarriages have been engineered. A
selection of them are examined and illustrated here.
There are no obvious rules on why some plants are more promiscuous than
others. Some of the most bizarre couplings unite species classified within
the same genus. A good example of this is Senecio 'Hippogriff (illustrated
in Cactus & Co. 6 (1): 48, 2002) combining S. articulatus and S. rowleyanus.
Here we are more concerned with those that bridge species of separate
genera, such as X Cremnosedum 'Little Gem' (Fig. 1), involving species of
Cremnophila and Sedum. The nothogeneric (hybrid generic) names aid in
pinpointing the ancestry. We call such examples intergeneric hybrids or
bigeners. Where several occur, we may doubt if the two "genera" are really
distinct, especially if the hybrids show some fertility: good pollen or seed
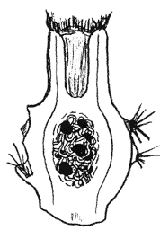
set. A good example is the cactus shown in Fig.2: a long-favoured basket
plant born in England in 1830 and attributed to Aporocactus flagelliformis
(the rat-tail cactus) X Heliocereus speciosus. Today both these genera are
amalgamated under Disocactus (Anderson 2001), so that what was an
intergeneric hybrid is now monogeneric: Disocactus X mallisonii. It has a
measure of self-fertility and sets some viable seed in small fleshy fruits
(Fig. 3). Most wide crosses, like the mule, are sterile. However,
intercrossability and intersterility are only one factor in determining the
distinctness of genera, and often it is more desirable to retain a pair of
genera as distinct despite the existence of crosses between them. In a
survey of succulent bigeners in 1982, the most numerous were classed under X
Pachyveria (Pachyphytum X Echeveria, 29 cases, Figs.4, 5) and X Gasteraloe
(Gasteria X Aloe, 24 cases, Figs.6, 7). Today there are doubtless more.

Fig. 11

Fig. 12

Fig. 13

Fig. 14

Fig. 15

Fig. 16

Fig. 17
| | | | | | |
Wide crosses are rare in nature (Fig. 8) because normally barriers have
arisen in the course of evolution. For instance, we find related species
flowering at different times, serviced by different pollinators or
genetically blocked at different stages from accepting alien pollen. In
cultivation it may be possible to overcome such barriers, and merely packing
plants together on a glasshouse staging may result in bizarre seedlings
coming up spontaneously. They can pose quite a problem when one tries to
guess the parentage.
All the largest families of succulents provide examples of wide crosses. It
used to be thought that the Aizoaceae (Mesembryanthemaceae) was an
exception, but this is no longer true. The breeding expertise of Steven
Hammer has resulted in many extraordinary wide crosses, 20 of which have
received nothogeneric names (Hammer 1995; Fig. 9)- In Cactaceae there are no
hybrids known between genera belonging to the different Subfamilies, and
some of the largest and most sharply defined genera of Cactoideae have
resisted efforts to cross their species with outsiders: Mammillaria,
Astrophytum, Ariocarpus, for example. Elsewhere the opposite is true, and
some remarkably dissimilar species have been successfully interbred. Fig. 10
shows Gil Tegelberg standing beside his plant raised by crossing the
red-flowered diurnal epiphyte Disocactus (Heliocereus) speciosus with the
columnar, nocturnal Pilosocereus palmeri. The extraordinary long-tubercled
Leuchtenbergia has been mated with species of Ferocactus and Thelocactus
with surprising results (Fig. 11). Differences in flower size and tube
length may not debar intercrossing, although when making the attempt it is
wise to use the short-tubed cactus as the female parent: its own pollen may
be unable to grow down the long style of the other. The longest flowers, as
in Echinopsis and Epiphyllum, are pollinated by hawk-moths; shorter narrow
tubes favour birds and the shortest funnel-shaped blooms bees. Thus in
nature these are kept apart by pollinator fidelity, but even here mistakes
can sometimes occur. In an analysis of 29 records of naturally occurring
cactus bigeners (Rowley 1994) I found about one third were between plants
adapted for different pollinators: bees, bats, birds or moths.
Fig. 12 shows typical flowers of two related genera: Gasteria with hanging,
tubular, red and yellow blooms typical of bird pollination, and Haworthia
with smaller, erect, oblique-limbed whitish flowers favoured by bees. The
hybrid between them (X Gasterhaworthia) has flowers of intermediate
character, but unlikely to suit either pollinator (Fig. 13). Hybrids
involving Haworthia X Aloe are surprisingly few. Fig. 14 shows one example,
as yet unflowered.
Some wide crosses are of curiosity value only. Echinocereus knippelianus X
Sclerocactus (Toumeya) papyracanthus (Fig. 15) sports a diversity of spines
from needle-like to flattened, weirdly disordered tubercles and small,
abortive flowers. A hybrid of Cleistocactus strausii X Echinocereus
(Wilcoxia) poselgeri (Fig. 16) is hardly an improvement on either parent. By
contrast, other wide crosses have great horticultural appeal combined with
hybrid vigour and ease of propagation. Such a one is Pachypodium 'Arid
Lands' (see title photo), a remarkable blend of two totally contrasted South
African species, P. namaquanum and P succulentum. Disocactus X mallisonii
(Fig.2) has already been mentioned; the North American and Mexican
Crassulaceae have spawned many desirable cultivars intermingling genes of
Echeveria, Graptopetalum, Pachyphytum and Sedum (Figs. 1, 4, 5, 17). Many
have arisen spontaneously and we can only guess the pedigrees, but they are
nonetheless worth adding to a collection. Who knows what further novelties
await the pollinator's brush?
REFERENCES
- ANDERSON, E. F. (2001). The Cactus Family. Timber Press, Oregon.
- HAMMER, S.A. (1995). New nothogenera, and a new combination in Mesembryanthema. Cact. Succ. J. (U.S.) 67: 172-173.
- ROWLEY, G.D. (1982). Intergeneric hybrids in succulents. Nat Cact. Succ. J. 37: 2-6, 45-49, 76-80, 119.
- ROWLEY, G.D. (1994). Spontaneous bigeneric hybrids in Cactaceae. Bradleya 12: 2-7.
Illustrations
Pachypodium 'Arid Lands', a wonderful creation from Chuck Hanson of Arid Lands Nursery in Arizona. P. succulentum (left) and P. namaqutanum (right) are the parents.
Fig. 1 Cremnosedum 'Little Gem', an intergeneric hybrid of Cremnophila nutans and Sedum humifusum
Fig. 2 Disocactus X mallisonii, a wide cross long favoured as a basket plant for conservatories
Fig. 3 Fruit in vertical section (x 4) of Disocactus X mallisonii showing a few plump seeds, It is 15 mm in diameter
Fig. 4 X Pachyveria pachyphytoides, a vigorous hybrid descended from Pachyphytum bracteosum X Echeveria gibbiflora Metallica'
Fig. 5 Another Pachyphytum bracteosum hybrid: X Pachyveria sobrina. The other parent is an unrecorded species of Echeveria
Fig. 6 Aloe jucunda (left), Gasteria batesiana (right) and the first generation hybrid between them, a handsome but unnamed X Gasteraloe.
Fig. 7 Another X Gasteraloe dating from 1896: X Gasteraloe lapaixii between its parents Gasteria bicolor (left) and Aloe aristata (right).
Fig. 8 X Hoodiapelia (Luckhoffia) beukmannii, a naturally occurring bigener attributed to Hoodia X Stapelia parentage.
Fig. 9 X Dinterops 'Stones Throw', Steven Hammer's cross of Dinteranthus vanzijlii X Lithops lesliei.
Fig. 10 Results of Gil Tegelberg's remarkable mating of Pilosocereus palmeri and Disocactus speciosus.
Fig. 11 Ferobergia 'Gil Tegelberg'. Crossing Leuchtenbergia principis with Ferocactus cylindraceus has lost some chlorophyll on the way.
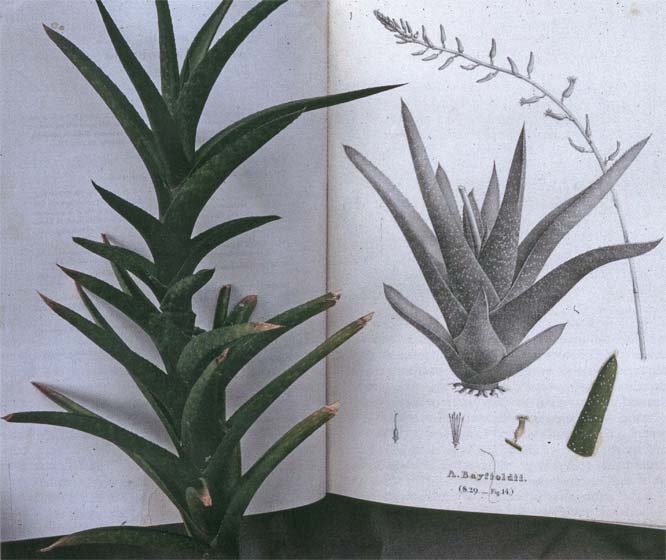
Fig. 12 Inflorescences of Gasteria (top), Haworthia (bottom) and a hybrid between them, X Gasterhaworthia bayfieldii.
Fig. 13 X Gasterhaworthia bayfieldii as illustrated by Salm Dyck in 1842, with the same sterile clone in cultivation today.
Fig. 14 A rare successful cross of Aloe X Haworthia: X Alworthia. The Aloe parent was A. bellatula, the Haworthia is unrecorded.
Fig. 15 A bit of everything inherited from Echinocereus knippelianus and Sclerocactus (Toumeya) papyracanthus. The flowers are all stillborn.
Fig. 16 Cleistocactus strausii X Echinocereus (Wilcoxia) poselgeri: another unexpectedly wide cross for the lover of oddities.
Fig. 17 Graptopetalum (Tacitus) bellum X Sedum suaveolens, a new X Graptosedum.