PUEBLOA, A NEW GENUS OF CACTACEAE JUSS.
Jornal "SUCCULENTS / СУККУЛЕНТЫ bilingual", 1999, 1(2), p.19-24
A. B. Doweld, National institute of Carpology (Gaertnerian institute)
ABSTRACT. The monotypic genus Puebloa Dwd. in the Cactaceae Juss. (P. bradyi (L. Bens.) Dwd.) is proposed to accommodate Pediocactus bradyi L. Bens. (P. winkleri Heil and P. despainii Goodr. & Welsh are treated as conspecific with P. bradyi), which is distinct from other species of the genus Pediocactus Br. & R. in having specific combination of tuberculate costate-sinuate or tuberculate-reticulate seed sculpturing and granular cuticular sculpturing, large stem sizes, lack of central spines, 12(14 ((15) radial spines, naked fruit dehiscing at maturity longitudinally and transversely. Puebloa is identified as the closest relative of Navajoa Croiz., on the basis of similar seed anatomy and morphology as well as similar morphology of flower, fruit, areoles, spination. From the evidence stated above, it is suggested that Puebloa seems represent a hypermorphic form of the navajoid ancestor. Puebloa together with allied genera Meyerocactus Dwd., Navajoa, Pediocactus Br. & R., and Utahia Br. & R. form a distinct evolutionary line diverging early from the very base of the tribe Cacteae, which is strongly supported by specific seed coat morphology; in consequence it is proposed to create a new subtribe Pediocactinae Dwd. of the tribe Cacteae to include all genera mentioned above.
 |
| Photo 1. Pediocactus-winkleri |
The genus Pediocactus established by N. L. Britton and J. N. Rose (in Britton & Brown, 1913) comprises usually 6 11 species, depending upon the authority (Benson, 1961/1962, 1982; Arp, 1972; Heil et al., 1981). Some of them are referred to as several distinct genera such as Navajoa Croiz. (P. peeblesianus (Croiz.) L. Bens.), Pilocanthus B. W. Bens. & Backbg. (P. paradinei B. W. Bens.) and Utahia Br. & R. (P. sileri (Engelm.) L. Bens.), and therefore Pediocactus s. str. comprises only 3 species (Backeberg, 1966): P. simpsonii (Engelm.) Br. & R., P. knowltonii L. Bens. and P. bradyi L. Bens. The last extensive revision of the genus was made by K. Heil et al. (1981), who confirmed the concept of L. Benson (1961/1962) of a polymorphic genus with the exception of Pediocactus papyracanthus (Engelm.) L. Bens., again excluded into a distinct genus Toumeya Br. & R. Nevertheless, the infrageneric system has not been re-considered since L. Benson (1961/1962), although its weak spots have been criticised by C. Backeberg (1966), who completely rejected it. The palynomorphological studies of B. Leuenberger (1976) also did not clarify the problem as to whether these taxa represent a single polymorphic genus, or a group of mostly monotypic genera.
 | | Photo 2. Pediocactus-winkleri |
K. Heil et al. (1981) published a series of the photomicrographs of Pediocactus seeds without any special comment. These researchers showed a great heterogeneity of seed morphology within polymorphic genus, which could not normally support the adopted taxonomic concept. Later G. Hentzschel (1990) presented a more detailed analysis of the seed morphology of Pediocactus and came to the conclusion that it is necessary to divide this polymorphic genus Pediocactus sensu L. Benson (1961/1962) into several natural genera. But, unfortunately, his conclusions have not been reflected in any of the new systems of the Cactaceae (Hunt, Taylor, 1990; Barthlott, Hunt, 1993; Eggli, Nyffeler, 1994). These students continued to treat the genus Pediocactus in its artificial volume in spite of the availability of other data precluding such a systematic treatment.
 |
| Photo 3. Navajoa-fa.maia |
The recent discovery of the unusual combination of the granular cuticular sculturing and tuberculate-sinuate seed sculturing with costal elements within Echinocactus horizonthalonius Lem., which has been separated into a distinct genus Meyerocactus Dwd. (Doweld, 1996), supports our former conclusions (Доуэльд, 1995) on the high systematic value of seed characters in the tribe Cacteae. The combination of the tuberculate types of seed sculturings and granular cuticular sculturing turned out to be very specific and occurred in this tribe only within the seeds of Navajoa, Utahia and Pediocactus bradyi-complex (incl. P. despainii and P. winkleri), while the other species of Pediocactus have a simple advanced papillate seed sculturing (Hentzschel, 1990; Доуэльд, 1995). The segregation of Echinocactus horizonthalonius into a distinct genus Meyerocactus on the basis of a quite distinct seed morphology has questioned the modern systematic status of P. bradyi-complex as a constituent part of the genus Pediocactus. These new findings and interpretations of the results of studies in seed morphology has forced us to re-examine in detail the evolution of seeds of P. bradyi-complex with reference to phylogeny and supposed origin of the whole 'pediocactoid' group of genera (Navajoa, Utahia, Pilocanthus) including a newly established ally Meyerocactus.
Materials and Methods
 |
| Photo 4. Pediocactus-despainii |
The mature seeds of Pediocactus bradyi L. Bens., P. winkleri K. Heil and P. despainii Goodr. & Welsh and other supposedly allied species (Table 1) were obtained from various field and horticultural seed collections; vouchers were deposited to the Carpotheca of the National Institute of Carpology (Gaertnerian Institution), Moscow [NICAR]. In addition to previous works on seed morphology of Pediocactus, carried out by K. Heil et al. (1981) and G. Hentzschel(1990), original Scanning Electron Microscope (SEM) observations were made with a HITACHI S-405 A at 15 kV after sputtercoating with platinum-palladium (O'Brien, McCully, 1981).
Table 1.
A summary of the principal characters differing between the genera of Pediocactinae
Characters / Taxa
Meyerocactus
Utahia
Navajoa
Puebloa
| Pediocactus | | | | | |
Stems
depressed-globose to ovoid or columnar 1015 (-50) cm long 10-15 cm wide
depressed-ovoid or ovoid 5-10 (-13) cm long 5-7.5 cm wide
obovoid, globose to depressed-globose 1.5-3.8 (-6.2) cm long 1.5-3.8 cm wide
subglobose to obovoid 3.8-6.2 cm long 2.5-5 cm wide
| depressed-ovoid to elongate-ovoid 2.5-12.5 (-30) cm long 2.5-10 (-12.5) cm wide
| | | | | |
Tubercles
coalescent into 7-13 ribs
truncate-conical
mammillate to truncate-pyramidal
cylidroid-ovoid or elongate-ovoid, ± truncate
| ± pyramidal
| | | | | |
Spines central
3-5
3-7
0-1
0 (1-2)
| 5-8 (-11)
| | | | | |
Spines radial
5-7
10-15
3-7
± 14-15
| 12-30
| | | | | |
Flower, diam. x long, cm
5-6.5 x 5-6.2
± 2.5 x ± 2
1.5-2.5 x 1.2
1.5-3 x 1.5-3
| 1.2-2.5 (-5) x 1.2-2.5 (-4)
| | | | | |
Fruit covered densely with hairs cylindroid
with several long-fimbriate scales cylindroid
smooth, with 1 or a few scales subcylindroid, broader above
smooth, basally constricted into short stalk broadly turbinate
| smooth, but with a few fimbriate scales cylindroidal
| | | | | |
Seed sizes, long x wide x thick, mm
4-4.8 x 2.5-3 x 2.5-3
(3.5-) 4.5-5 x (3-) 3.5-4.3 x 2.5-3.5
2.5-3 (-3.5) x 2-2.5 x 2-2.5
2.5-3.5 x 1.8-2.5 x 1.5-2.5
| (1.5-) 2.5-3 (-3.5) x (1.5-) 2-3 x 0.9-2.1
| | | | | |
seed sculpture
tuberculate-sinuate with costal elements
tuberculate-sinuate
tuberculate-reticulate with costate-reticulate elements (if any)
tuberculate costate-sinuate to tuberculate-reticulate with costal elements
| papillate
| | | | | |
cuticular sculpture
granular
granular or diffusive reticulate-granular
granular or reticulate-granular
granular or enlarged granular
| granular
| | | | | |
Additional observations on seed anatomy and morphology of P. bradyi-complex
 |
| Photo 5. Pediocactus-bradyi |
Seeds (Figs. 1, 2, 5, 8, 9) are obliquely obovoid; 2.5 3.5 mm long, 1.8 2.5 mm broad, 1.5 2.5 mm thick; dark brown to black. The hilum (Figs. 3, 6, 10) is smooth and concaved, rectangular; vascular bundle track protruding; micropyle distinct from the hilum, slit-shaped or elliptic, depressed by overmicropylar rostrum turning into a 4 6-rowed antiraphe. Seed sculturing is tuberculate costate-sinuate (Figs. 1, 9) to largely tuberculate-reticulate (Fig. 5, 8) with costal elements. Cuticular sculturing is granular or enlarged granular (Figs. 4, 7, 11).
The spermoderm is 4 5-layered (Fig. 12), originated from both integuments (outer integument: 3 layers, inner integument: 2 3), exotestal-endotegmic. The exotestal cells (solitary layer derivative of the outer epidermis of outer integument) are large and radially oblong, cupolashaped, (rectangular in cross-section. Their outer walls are thickened and traversed by numerous microchannels, which are ended by tuberculate structures forming granular cuticular sculturing. The 1-layered endotegmen is composed of (well developed (only locally depressed) cells with a few-thickened walls; the cells are filled with tannins (brown). Other inner layers of the seed coat (2 3) are thin-walled and mostly obliterated.
 |
| Photo 6. Pediocactus-bradyi |
P. bradyi L. Bens. Seeds are 2.5 3 ( 3.5, in the southern populations!) mm long, 1.8 2.2 ( 2.5, in the same places) mm broad, 1.5 1.8 ( 2, in the same places) mm thick. Seed sculturing is [tuberculate] costate-sinuate in the northern part of area (Fig. 1), but southward the 'sinuate costality' turns into (tuberculate-reticulate seed sculturing (Fig. 2).
P. winkleri Heil. Seeds are 3 3.5 mm long, 2 2.5 mm broad, 1.8 2.5 mm thick. Seed sculturing (Fig. 5) is (largely tuberculate-reticulate or with costal elements (mostly in the northern part of area).
P. despainii Goodr. & Welsh. Seeds are 3 3.5 mm long, 2 2.5 mm broad, 1.8 2.5 mm thick. Seed sculturing is varying from tuberculate-reticulate with costal elements (Fig. 8) to [tuberculate] costate-sinuate (Fig. 9) in the northern part of area.
Discussion
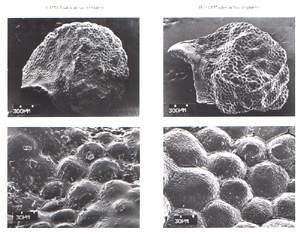 |
| Photo 7. 070-P.winkleri-010-P.despai |
Relationships of Pediocactus bradyi-complex with miscellaneous genera. The P. bradyi-complex having an unusual combination of the tuberculate costate-sinuate seed sculturing and granular cuticular sculturing is sharply distinct from other species of Pediocactus s.str. [P. simpsonii (Figs. 13-14), P. paradinei (Fig. 15-16) and P. knowltonii (Fig. 17-18)], which are characterized by simple papillate seeds lacking in tuberculate specialisation of exotesta. However, P. bradyi-complex is similar to such distinct genera as Navajoa (having a tuberculate-reticulate seed sculturing: Figs. 19-20), Utahia (tuberculate-sinuate seed sculturing: Figs. 21-22) and Meyerocactus (tuberculate-sinuate: Doweld, 1996). The similarity of P. bradyi-complex with Navajoa is supplemented by the occurrence of similar tuberculate-reticulate seed sculturings within some populations of P. winkleri and P. despainii p.p. Taking into consideration a striking resemblance of P. bradyi-complex with distinct genera Navajoa, Utahia and newly established Meyerocactus in seed and cuticular sculturing, and striking differences (table 2) in the form of tubercles, larger stem sizes, lack of central spines, distinctive arrangement of 12 14 ( 15) radial spines, naked fruit (Benson, 1961/1962, 1982), and at the same time noticing a full dissimilarity with other species of Pediocactus s. str. (including type-species, P. simpsonii, having papillate seeds), we think it necessary (as it formerly has been stated provisionally by G. Hentzschel, 1990) to exclude P. bradyi-complex from the genus Pediocactus into a distinct genus Puebloa.
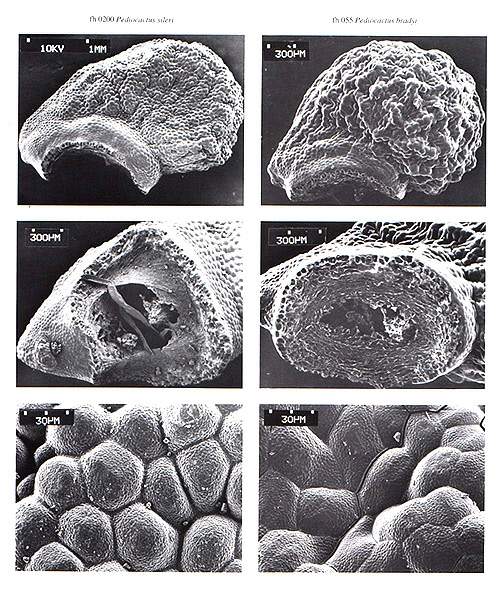 |
| Photo 8. 0200-P.sileri-055-P.bradyi |
Puebloa A. Doweld, gen. nov. Plantae globosae vel applanato-globosae, 3.8 6.2 cm lg., 2.5 5 cm diam., areolae elliptici, aculei radiali 12 14 ( 15), centrali solitarii vel absentis, fructus nudus, baccatus, maturus exsiccans, longitudinale et circumscissile dehiscens, 1 cm lg., 1.5 cm diam., semina nigra vel obscure brunea, porus micropylario distinctus, exotesta tuberculata costato-flexuosa vel tuberculato-reticulata, embryo curvatus, perispermium magnum.
T y p u s: Puebloa bradyi (L. Bens.) A. Doweld ( Pediocactus bradyi L. Bens.
The genus is named in the honour of the Pueblo people, living in the areas of the genus distribution.
The creation of the new genus Puebloa is the next step in the revisionary studies of the whole tribe Cacteae towards a great understanding of its origin, phylogeny and systematics. The seed anatomy and morphology confirms the status of distinct monotypic genera Utahia, Navajoa, Pediocactus s. str. and newly established Puebloa and refutes the former conclusions of W. Barthlott (1988) on the unity of these genera under Pediocactus, who did not document them by photomaterials.
The unusual similarity of seed sculpture's types within Puebloa bradyi ssp. winkleri (incl. both varieties winkleri and despainii) with the seeds of Navajoa, normally indicative of very close phylogenetic relationships (Доуэльд, 1995), should be explained. The species of Puebloa and Navajoa are sharply distinct in habitus: the stems of Puebloa are larger and more robust than that of Navajoa (3.8 6.2 x 2.5 5 cm vs. 1.5 3.8 x 1.5 3.8 cm), tubercles in Puebloa are elongate-ovoid (truncate-cylindroidal), but in Navajoa they are mamillate or truncate-pyramidal. At the same time there is a striking resemblance between two genera in the general construction of areole and spination: in both genera the central spines are wanting or rarely 1 2; the number of radial spines in Navajoa is (3 ) 6 7, in contrast to Puebloa, the radials of which are represented in duplicate number: 12 14 ( 15). In both genera the ovary are normally without scales (if any, 1 or a few on the upper portion of fruits). The striking likeness in seed and fruit morphology, and general construction of areole and spination between Puebloa and Navajoa are undoubtedly indicative of close phylogenetic relationships. Such a relationship might be explained, if we assume, judging from gross sizes of Puebloa stems, that Puebloa is a peramorphic derivative of one of the Navajoa species, showing, perhaps, a hypermorphic state of its morphology (de Beer, 1930; Alberch et al., 1979; McNamara, 1986). The extension of comparatively late ontogenetic development which is characteristic for hypermorphosis, may result in morphological characteristics quite different from the ancestral ('navajoid') adult: (1) in the attainment of larger sizes (cf. stem sizes for Puebloa), and (2) the hypermorphic form will continue developing into next morphological stage of specialisation (i.e. duplication of radial spines in Puebloa). This putative scenario could explain the unexpected nearness of two genera, Navajoa and Puebloa, the last of which might represent a hypermorphic form of the navajoid ancestor.
The specific combination of various tuberculate seed sculturing types and granular cuticular types, occurring only in the whole tribe Cacteae within Utahia, Navajoa, Puebloa and Meyerocactus, indicates the early divergence of this evolutionary line from the ancestral complex of the tribe. The resemblance of seed sculturings in Pediocactus-like genera (usually papillate) and in Sclerocactus-like genera (also papillate) might be considered to be proof of the close phylogenetic relationships between these groups of genera as has been suggested by F. Buxbaum (1951 : 55-56). His phylogenetic 'Seitenlinie Pediocactus Utahia' was considered to be a direct derivative of the Sclerocactus-line (subtr. Sclerocactinae Dwd.), the primitive state of which is currently confirmed in addition by the complete lack of cuticular sculturing and occurrence of papillate seed sculturing (Doweld, 1996; Доуэльд, 1998). The morphogenetic hiatus between the pediocactoid seed type (typically papillate seed sculturing plus granular cuticular sculturing) and the sclerocactoid seed type (papillate/apapillate seed sculturing and smooth cuticular sculturing: Figs. 23-24) is so great that it is impossible to agree with inclusion or even nearness of sclerocactoid Toumeya (having finely papillate seed sculturing and lacking of cuticular sculturing: Figs. 25-26) to the genera of Pediocactus-group as has been made by L. Benson (1961/1962, 1982). At the same time the proposed relationships of Navajoa with Toumeya (Buxbaum, 1951) sensu H. Bravo and W. Marshall (1956, 1957), including several species currently recognized in Turbinicarpus (Kladiwa, 1963, 1965), have also turned out to be artificial: species of Turbinicarpus have a distinctive papillate seed sculturing with a striate cuticular sculturing (Figs. 27-28). This combination is quite distinct in morphogenetic origin and has nothing in common with the appearance of granular cuticular coat within papillate seeds (Доуэльд, 1995); that is reflected in the placement of Turbinicarpus into a distinct subtribe Turbinicarpinae Dwd. (Доуэльд, 1998), rooting in the archaic Sclerocactinae and Bravocactinae Dwd.
Relationships of Puebloa with subtr. Thelocactinae.
Subtr. Pediocactinae
The genera of the Pediocactus-group reveal a certain relationship with the genera of the subtribe Thelocactinae F. Buxb. emend. Dwd. such as Thelocactus, Neolloydia, Echinofossulocactus, Leuchtenbergia, and Obregonia (Доуэльд, 1995). They are also characterized by papillate seed sculturing with granular cuticular sculturing, but with faceted thickenesses on the cell borders. This so-called faceted-granular cuticular sculturing is more specialised than a simple granular cuticular sculturing of pediocactoid seeds, and it might be indicative of a certain advance of the Thelocactinae, which is supplemented by their specialised structure of the hilum-micropylar region and seed form. The pediocactoid genera Meyerocactus, Utahia, Navajoa, and Puebloa are specialised in quite another direction, having advanced tuberculate seed sculturings originated on the basis of a papillate type. With the segregation of Pediocactus bradyi-complex into a distinct genus Puebloa having tuberculate-reticulate (sinuate) seed sculturing, the amplified genus Pediocactus became a more natural and uniform (in having only simple papillate seeds), and thus may serve a connecting link to advanced Thelocactinae with similar papillate seeds covered with specialized faceted-granular cuticular sculpturings. Nevertheless, all genera of the Pediocactus group possess a more primitive, simple granular cuticular sculturing and structure of the hilum-micropylar zone of seed (without 'hilar cup'). These essential differences preclude any possibility to consider them as part of the subtribe Thelocactinae (Buxbaum, 1958).
The essential differences in seed structure of the genera of the subtribe Echinocactinae, formerly discussed in detail by F. Buxbaum (1951) and A. Doweld (1996; Доуэльд, в печати), do not support their inclusion in this subtribe as well as in any other existing subtribes of Cacteae (Доуэльд, 1998). In our opinion, the specific combination of the tuberculate seed sculturings with granular cuticular sculturings among the distinctive pediocactoid genera of the whole tribe Cacteae is indicative of the necessity to create a new distinct subtribe Pediocactinae Dwd. to accommodate these anomalous group of taxa (Table 3). The creation of the new subtribe opens new perspectives in the development of a new detailed natural phylogenetic system of the tribe Cacteae with clearly defined generic and suprageneric boundaries.
The infraspecific system of Puebloa
Seed morphology of the Pediocactus bradyi-complex is indicative of the unusual nearness of these three species constituting a very specialised group. The similarity in seed sizes and the intertransitority of seed sculturings within Pediocactus winkleri and Pediocactus despainii are indicative, in our opinion, of the incompetence of the elevation of the races into species. A somewhat unusual geographical transition from the type of seed sculturing to another, more specialized (tuberculate-reticulate to [tuberculate] costate-sinuate) within populations of P. bradyi and their geographical distribution within P. winkleri and P. despainii could indicate that these three species are conspecific (a single polymorphic species, P. bradyi). Judging from seed morphological data now available, we agree with F. Hochstдtter (1994), who united for the first time all three species under Pediocactus bradyi But from the view point of seed morphology it seems to us that it is expedient to consider P. winkleri and P. despainii as varieties only of a single subspecies (winkleri), constituting thus a more extensive northern geographical race, and quite distinct from the southern geographical race concluding P. bradyi. With the creation of a new genus Puebloa all discussed species are transferred to a new genus and united under a single species, Puebloa bradyi (L. Bens.) Dwd.
Puebloa bradyi (L. Bens.) A. Doweld, comb. nov. Pediocactus bradyi L. Bens., 1962, Cact. Succ. J. (Los Angeles) 34, 1 : 19; ejusd., 1982, Cacti U.S. Can. : 756, p.p.; Hochst., 1994, Succulenta 73, 1 : 55.
T y p u s: Near the Marble Canyon of the Colorado River, Coconino Co., northern Arizona, 21 IV 1961, L. & E. L. Benson, n( 16807 (POM).
1. Puebloa bradyi subsp. bradyi Toumeya bradyi (L. Bens.) W. H. Earle, 1963, Cacti SW. : 97. Pilocanthus bradyi Glass et Foster, 1973, Cact. Succ. J. (Los Angeles) 45 : 254, nom. nud.
T y p u s: as in species.
D i s t r i b u t i o n: Area near Marble Canyon of the Colorado River, Coconino Co., Arizona, U.S.A.
2. Puebloa bradyi subsp. winkleri (Heil) A. Doweld, comb. et stat. nov. Pediocactus winkleri Heil, 1979, Cact. Succ. J. (Los Angeles) 51, 1 : 25-30. Pediocactus bradyi var. winkleri (Heil) Hochst. 1994, Succulenta 73, 2 : 55. Pediocactus bradyi ssp. winkleri (Heil) Hochst. 1995, Gen. Pediocactus, Navajoa, Toumeya Rev. : 61.
T y p u s: U.S.A., Utah, Wayne Co., 5 IV 1977, K. D. Heil, n( 64285 (UNM).
2a. Puebloa bradyi ssp. winkleri var. winkleri
T y p u s: as in subspecies.
Di s t r i b u t i o n: U.S.A., Southeastern (Wayne Co.) Utah, in the Navajoan Desert at 1500 1600 m elev.
2b. Puebloa bradyi ssp. winkleri var. despainii (Welsh & Goodr.) A. Doweld, comb. et stat. nov. Pediocactus despainii Welsh & Goodr. 1980, Great Bas. Nat. 40, 1 : 83-86. Pediocactus bradyi var. despainii (Welsh & Goodr.) Hochst. 1994, Succulenta 73, 2 : 57. Pediocactus bradyi ssp. despainii (Welsh & Goodr.) Hochst. 1995, Gen. Pediocactus, Navajoa, Toumeya Rev. : 61.
T y p u s: U.S.A., Utah, Emery Co., San Rafael Swell, 5 V 1978, K. Despain, n( 266A (BRY).
D i s t r i b u t i o n: U.S.A., Utah, Emery Co., San Rafael Swell (so far as known).
Acknowledgements
The author is obliged to Mr. S. Brack, Mr. J. Busek, Mr. M. Hajek, Mr. F. Hochstдtter, Mr. E. Lutz, Mr. D. Rowland, Mr. J. Schraets for providing documented materials for present study. Special thanks go to Mrs. Francesca B. Yakovleva for invaluable technical assistance with SEM studies.
LITERATURE CITED
- Д о у э л ь д А. Б. Анатомия, морфология и ультраструктура семенных покровов трибы Cacteae в связи с её систематикой и филогенией. Т. 1 2. М., 1995. 156, 280 с.
- Д о у э л ь д А. Б. Конспект филогенетической системы трибы Cacteae (Cactoideae-Cactaceae). I. Система подтриб Sclerocactinae-Bravocactinae-Turbinicarpinae // Суккуленты. 1998. № 1. С. 15-30.
- Д о у э л ь д А. Б. Обзор трибы Cacteae. Роды и надродовые таксоны, 1 // Новости систематики высших растений. T. 32. CПб, 1999.
- A l b e r c h P., G o u l d S. J., O s t e r G. F., W a k e D. B. Size and shape in ontogeny and phylogeny // Paleobiology. 1979. Vol. 5, № 3. P. 296-317.
- A r p G. A revision of Pediocactus // Cact. Succ. J. (Los Angeles). 1972. Vol. 44. № 4. P. 218-222.
- B a c k e b e r g C. Das Kakteenlexikon. Jena, 1966. 741 S.
- B a r t h l o t t W. Ьber die systematische Gliederungen der Cactaceae // Beitr. Biol. Pflanz. 1988. Bd. 63. H. 1. S. 17-40.
- B a r t h l o t t W., H u n t D. R. Cactaceae // The families and genera of flowering plants / Ed. K. Kubitzki. Vol. 2. Berlin—Wien, 1993. P. 161-197.
- B e e r G. R. de. Embryology and Evolution. Oxford, 1940. 116 p.
- B e n s o n L. A revision and amplification of Pediocactus // Cact. Succ. J. (Los Angeles). 1961/1962. Vol. 33, № 1. P. 49-54; Vol. 34, № 1, 3, 4. P. 17-19, 57-61, 163-168.
- B e n s o n L. The Cacti of the United States and Canada. Stanford, 1982. 1044 p.B r a v o H. H., M a r s h a l l W. T. A revision of the genus Toumeya // Saguaroland Bull. 1956. Vol. 10. P. 112-119.
- B r a v o H. H., M a r s h a l l W. T. The genus Toumeya // Saguaroland Bull. 1957. Vol. 11. № 1. P. 31.
- B r i t t o n N. L., B r o w n A. An illustrated flora of the northern United States and Canada, and the British possessions: from Newfoundland to the parallel of the southern boundary of Virginia, and from the Atlantic Ocean westward to the 102d meridian. 2nd ed. Vol. 2. Amaranthaceae Loganiaceae. Amaranth to Polyprenum. New York, 1913. 735 p.
- B u x b a u m F. Die phylogenie der nordamerikanischen Echinocacteen // Цsterr. Bot. Zeitschr. 1951. Bd. 98, H. 1. S. 44-104.
- B u x b a u m F. The phylogenetic division of the subfamily Cereoideae, Cactaceae // Madroсo. 1958. Vol. 14. P. 177-206.
- D o w e l d A. B. De polyfylie van het geslacht Echinocactus aangetoond door zaadhuidstructuur en -ornamentatie // Succulenta. 1996. Vol. 75, № 6. P. 268-278.
- H e i l K., A r m s t r o n g B., S c h l e s e r D. A review of the genus Pediocactus // Cact. Succ. J. (Los Angeles). 1981. Vol. 53, № 1. P. 17-39.
- H e n t z s c h e l G. Morphology of the seeds of Pediocactus and Sclerocactus species // F. Hochstдtter. To the habitats of Pediocactus and Sclerocactus. Mannheim, 1990. P. 134-150.
- H o c h s t д t t e r F. Pediocactus bradyi, een revisie 2 // Succulenta. 1994. Vol. 73, № 2. P. 55-62.
- H u n t D. R., T a y l o r N. P. The genera of Cactaceae: progress towards consensus // Bradleya. 1990. Vol. 8. P. 85-107.
- K l a d i w a L. Das Toumeya-Problem und die eingezogenen Genera Navajoa und Turbinicarpus // Sukkulentenkunde. 1963. Bd. 7/8. S. 84-86.
- K l a d i w a L. Das Genus Toumeya, seine Arten und Variabilitдt // I.O.S. Bull. 1965. Vol. 2, № 3. P. 84-86.
- L e u e n b e r g e r B. E. Die Pollenmorphologie der Cactaceae und ihre Bedeutung fьr die Systematik. Mit Beitrдgen zur Methodik rasterelektronenmikroskopischer Pollenuntersuchungen und mit EDV-unterstьtzten Korrelationsanalysen. Vaduz, 1976. 321 S. (Diss. Bot., 31).
- M c N a m a r a K. J. A guide to the nomenclature of heterochrony // Jour. Paleontol. 1986. Vol. 60, № 1. P. 4-13.
- N y f f e l e r R., E g g l i U. Die Konsensusklassifikation der Kakteen // Kakt. and. Sukk. 1994. Bd. 45. S. 142-149.
- O ' B r i e n T. P., M c C u l l y M. E. The Study of Plant Structure: Principles and selected Methods. Melbourne, 1981. 290 p.
Photos:
Photo 1 - Photos from the book by Edward F. Anderson "The cactus family" – 2001
Photo 3 – Photos from I-net
Photo 2, 4 – 8 – Photos from the book by Fritz Hochstatter "An den Standorten von Pedio- und Sclerocactus" - 1989
Photos from the book were copied by Oleg Pikalev, Moscow, URL: http://www.cactuslove.ru